 subscribe to horse genetics
subscribe to horse genetics




horse genetics
 Ever wondered just what horse genetics really is? If so visit the "What is horse genetics" page to find out! Stay here to learn about the basic principles of genetics as applied to horses. In addition to the text and diagrams I will now be incorporating more photos, and videos, to illustrate and help clarify some of the principles and methods presented. So keep coming back to learn and refresh your knowledge! Ever wondered just what horse genetics really is? If so visit the "What is horse genetics" page to find out! Stay here to learn about the basic principles of genetics as applied to horses. In addition to the text and diagrams I will now be incorporating more photos, and videos, to illustrate and help clarify some of the principles and methods presented. So keep coming back to learn and refresh your knowledge!
Understanding basic horse genetics is the key to understanding horse color and coat patterns, as well as the genetic disorders of the horse, such as lethal white syndrome. Outside of horse breeding genetics is also important for other reasons, for example conservationists now use a knowledge of horse genetics to help conserve the Przewalski's and other endangered horse populations.
For most readers this section can be used as a reference, to fill in your basic knowledge of horse genetics as required. Only students on a formal genetics course ought to know all of what is presented here. They should also realise, however, that this is not a comprehensive coverage of basic genetics, but just enough to understand what is presented on this web-site.
This page covers various aspects of essential horse genetics basics including about genes, alleles, chromosomes, Mendelian inheritance for one and two characters and modified ratios, partial dominance, co-dominance, lethal alleles, epistasis, genetic linkage, partial linkage (example) and sex-linkage. There is also a section with horse genetics references and further reading. Happy reading! 
|

|
| To understand horse genetics you need to know what genes are Anyone who wants to understand something about horse genetics should have some idea of what genes are and what they do, even if it’s only at an elementary level.
Genes are the units of hereditary. They can’t be seen directly, but the results of their action can be seen in all of life, including horses.
Genes are like pieces of code that say how the molecular building blocks of living organisms will be built, what will be put or done where, and when. This in turn determines what those organisms look like and how they work (or don’t). The genes are encoded by a molecule called DNA (deoxyribonucleic acid). Since Watson and Crick discovered the structure of DNA in 1953 there has been an explosion of knowledge that has revolutionised genetics, including horse genetics. As a consequence we now have quite a deep understanding of what genes are and how they work. Horse genetics is now being studied at the level of gene structure and function, leading to an increased understanding of both genetic disorders and color inheritance.
According to the human genome project humans have about 30,000 genes (18/2/2001). This is only a few thousand more than mice are thought to have, and one guesses that horses have about the same number of genes as us. With the exception of some very special cells, which are involved in making our immune systems work, each of our cells contains all of the 30,000 or so genes - the same is true of horses. Since (almost) all body cells inherit the same genetic information we might ask why they aren’t all identical to one another. In general the reason is to do with the way in which the different parts of the genetic information are used (or not used) in different cells.
Another branch of horse genetics is more concerned with the way genes work, or don’t work. Research in this area of horse genetics leads to an increased understanding of genetic disorders and can lead to genetic tests for identifying carriers, as well as to new or improved methods of managing or treating disorders in known sufferers. |
|

Genes occur in different forms called alleles
Alleles are gene variants caused by mutation. It is important not to confuse genes and alleles. It is one of the commonest mistakes among amateur geneticists and frequently causes confusion among horse breeders. I suggest reading the text here AND watching the video! Each allele has a slightly different code and may make a slightly different product, or control a process in a slightly different way. It is rather like models of a particular make of car, in that each model is essentially similar with just some minor differences. (Hopefully!) each model of car will transport you around wherever you want to go, but some will do it faster, some will be more comfortable, or look sleeker, while others will have greater fuel efficiency, and yet others will be better for particular types of terrain.
The above description of natural selection is simplified of course. The observant reader might well ask why we do not eventually end up with only perfect organisms with no genetic disorders or defects. In fact the reasons are various, and are now well known and researched. One is that some mutations have more than one effect, or have a different effects in different environments, so that evolution is both chucking them away in one place or time and preserving them in another. Another reason is that some recessive mutations hide in heterozygotes, which are explained later.
If an allele has to be homozygous to affect the phenotype it is said to be recessive, an example is the red allele of the Extension gene - chestnut horses are homozygous for the recessive allele.
If an allele over-rides another, as the grey allele overides the non-grey allele, for example – the phenotype is affected even when the allele is heterozygous - the allele is said to be dominant.
|



Genes are organised on chromosomesGenes don't just float around in cells. They are spaced out along a (relatively) few long continuous strands of DNA. Often the analogy of beads on a string is used. This is quite a good analogy, though the beads aren't spaced out evenly: sometimes there are clusters, sometimes there are long spaces between the genes. Furthermore we now know that there's much more string than beads, with much DNA not coding for anything at all!
The DNA strands are very long and are folded, coiled and held together in a way that makes up structures called chromosomes. These can be fitted into cells without becoming tangled and broken. They help to ensure that hereditary takes place without making too many mistakes that might otherwise result in abnormal and inviable cells and offspring.
Horse cells have 64 chromosomes each, located in a sub-cellular compartment called the nucleus. Particular places on a chromosome are called loci (singular locus). Each particular gene has a particular place, or locus, on a particular chromosome. A particular gene is always located at the same locus.
Each cell of a horses body contains two copies of each chromosome - one from the dam and one from the sire. In horses there are 32 pairs of chromosomes. The paternal and maternal chromosome of each pair are said to be homologous. This means that they have same genes, and other structural features, at corresponding places along their length (and they are therefore structurally the same).
An example of a sex-linked gene is that causing haemophilia A (factor VIII deficiency). Although the example is not unique to horse genetics, it has been reported rarely in Thoroughbred, Quarter Horse and Standardbred colts (Archer, 1961, Henninger, 1988, Hutchins et al, 1967).
|

_____







 We can demonstrate the affect of epistasis on genetic ratios through a horse genetics example. Say we had a bay mare and stallion who were both of genotype E+e at the extension locus (E+ causes the production of the black eumelanin pigment) and of genotype AAAa at the agouti locus (which controls the distribution of black pigment).
We can demonstrate the affect of epistasis on genetic ratios through a horse genetics example. Say we had a bay mare and stallion who were both of genotype E+e at the extension locus (E+ causes the production of the black eumelanin pigment) and of genotype AAAa at the agouti locus (which controls the distribution of black pigment). 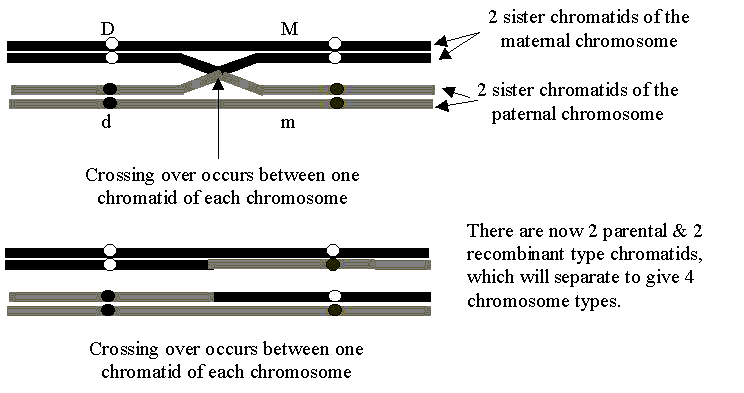
 The extension coat color gene (E locus) is linked to the genes for roan (RN) and tobiano (To). The dominant extension allele is E+ and horses with this allele produce black eumelanin pigment, resulting in the horses of black, brown or bay and their derivatives, depending on other genes. Allele e is recessive and horses of genotype ee are chestnut (including sorrel) or some derivative of that color (such as palomino). The dominant allele at the tobiano locus is ToT, with horses of genotype To+To+ being solid colored rather than white spotted.
The extension coat color gene (E locus) is linked to the genes for roan (RN) and tobiano (To). The dominant extension allele is E+ and horses with this allele produce black eumelanin pigment, resulting in the horses of black, brown or bay and their derivatives, depending on other genes. Allele e is recessive and horses of genotype ee are chestnut (including sorrel) or some derivative of that color (such as palomino). The dominant allele at the tobiano locus is ToT, with horses of genotype To+To+ being solid colored rather than white spotted.


